................................................................. Biodiversity of plankton by species oscillations and chaos Jef Huisman *†‡ & Franz J. Weissing § of * Biological Sciences, Stanford University, Stanford, California 94305-5020, USA † Center for Estuarine and Marine Ecology, CEMO-NIOO, PO Box 140, 4400 AC Yerseke, The Netherlands § Department of Genetics, University of Groningen, PO Box 14, 9750 AA Haren, .............................................................................................................................................. The Netherlands Biodiversity has both fascinated and puzzled biologists 1 . In aqua- .............................................................................................................................................. tic ecosystems, the biodiversity puzzle is particularly trouble- NATURE | VOL 402 | 25 NOVEMBER 1999 | www.nature.com some, and known as the ‘paradox of the plankton’ 2 . Competition theory predicts that, at equilibrium, the number of coexisting species cannot exceed the number of limiting resources 3–6 . For phytoplankton, only a few resources are potentially limiting: nitrogen, phosphorus, silicon, iron, light, inorganic carbon, and sometimes a few trace metals or vitamins. However, in natural waters dozens of phytoplankton species coexist 2 . Here we offer a solution to the plankton paradox. First, we show that resource competition models 6–10 can generate oscillations and chaos when species compete for three or more resources. Second, we show that these oscillations and chaotic fluctuations in species abundances allow the coexistence of many species on a handful of resources. This model of planktonic biodiversity may be broadly applicable to the biodiversity of many ecosystems. We consider a well-known resource competition model 6–10 that
d N i d t ¼ N i ð m i ð R 1 ; … ; R k Þ � m i Þ i ¼ 1 ; … ; n ð 1 Þ n d R j d t ¼ D ð S j � R j Þ � � c ji m i ð R 1 ; … ; R k Þ N i j ¼ 1 ; … ; k ð 2 Þ i ¼ 1 … � � r i R 1 r i R k m i ð R 1 ; … ; R k Þ ¼ min ; … ; ð 3 Þ K 1 i þ R 1 K ki þ R k � � 1 : 00 0 : 90 0 : 30 1 : 04 0 : 34 0 : 77 0 : 30 1 : 00 0 : 90 0 : 71 1 : 02 0 : 76 K ¼ � � � � 0 : 90 0 : 30 1 : 00 0 : 46 0 : 34 1 : 07 r i ¼ 1 d � 1 and m i ¼ D ¼ 0 : 25 d � 1 for 8,11–16 � � 0 : 04 0 : 07 0 : 04 0 : 10 0 : 03 0 : 02 0 : 08 0 : 08 0 : 10 0 : 10 0 : 05 0 : 17 C ¼ � � � � 0 : 14 0 : 10 0 : 10 0 : 16 0 : 06 0 : 14
c 70 a 50 60 Species abundances 1 50 40 Species abundances 1 2 3 5 40 30 30 2 20 20 3 10 4 6 0 0 3,000 6,000 9,000 12,000 15,000 10 Time (days) d 0 150 200 0 50 100 40 Time (days) Species abundances 30 b 50 20 40 10 Species 3 30 0 0 600 1,200 1,800 2,400 3,000 20 Time (days) 10 0 10 20 Species 1 Figure 1 Oscillations on three resources. a , Time course of the abundances of three 30 0 10 40 20 species competing for three resources. b , The corresponding limit cycle. c , Small- 30 50 40 50 amplitude oscillations of six species on three resources. d , Large-amplitude oscillations of Species 2 nine species on three resources.
b Species 5 55 50 25 45 30 35 40 a 20 Species abundances Species 3 25 20 10 30 40 50 60 0 30 0 35 40 18 15 S p 100 e 12 c i e 9 s 1 6 Time (days) Total biomass c 120 150 60 90 30 0 0 4 200 2 1 5 3 100 Time (days) 300 200 300 Figure 2 Chaos on five resources. a , Time course of the abundances of five species competing for five resources. b , The corresponding chaotic attractor. The trajectory is plotted for three of the five species, for the period from t ¼ 1 ; 000 to t ¼ 2 ; 000 days. c , Time course of total community biomass.
Figure 3 Bifurcation diagram, for five species competing for five resources. The graphs show the local minima and maxima of species 1, plotted during the period from t ¼ 2 ; 000 to t ¼ 4 ; 000 days, as a function of the half-saturation constant K 41 . Part of a is magnified in b .
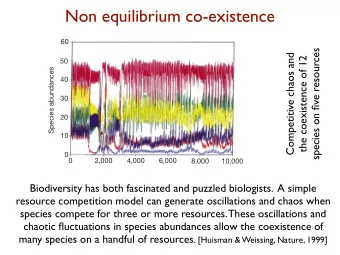
a b Species abundances Species abundances 20 12 16 20 30 40 50 10 60 8 0 4 0 0 0 2,000 2,000 4,000 4,000 Time (days) 6,000 6,000 8,000 8,000 10,000 10,000 Figure 4 Competitive chaos and the coexistence of 12 species on five resources. a , The abundances of species 1–6; b , the abundances of species 7–12.
(Fig. 4). The possibility that competition models may generate oscilla- tions and chaos was already recognized in the mid 1970s 28–30 . Also, it is well established that non-equilibrium conditions may favour species coexistence 5,12,20 . What is new here is that we found both phenomena in a single competition model. Moreover, our findings do not stem from an artificially constructed model, but are based on one of the standard models of phytoplankton competition 6–16 . We conclude that the biodiversity of plankton communities need not be
Vol 461 j 3 September 2009 j doi:10.1038/nature08227 REVIEWS Early-warning signals for critical transitions Marten Scheffer 1 , Jordi Bascompte 2 , William A. Brock 3 , Victor Brovkin 5 , Stephen R. Carpenter 4 , Vasilis Dakos 1 , Hermann Held 6 , Egbert H. van Nes 1 , Max Rietkerk 7 & George Sugihara 8 Complex dynamical systems, ranging from ecosystems to financial markets and the climate, can have tipping points at which a sudden shift to a contrasting dynamical regime may occur. Although predicting such critical points before they are reached is extremely difficult, work in different scientific fields is now suggesting the existence of generic early-warning signals that may indicate for a wide class of systems if a critical threshold is approaching.
a b Small forcing Large external change System state System state Across a non-catastrophic Affecting an almost threshold linearly responding system Conditions Conditions c d Small forcing Small forcing System state System state Across a Across the border of F 2 F 2 catastrophic bifurcation a basin of attraction F 1 F 1 Conditions Conditions Box 1 j Critical transitions in the fold catastrophe model
Low resilience High resilience d a Basin of attraction Basin of attraction Potential Potential High recovery rate Low recovery rate Disturbances Disturbances 0 2 4 6 8 10 0 2 4 6 8 10 State State 7.8 5.95 5.95 7.8 f c e b 7.76 5.9 5.9 7.75 State t +1 7.72 State t +1 State State 5.85 5.85 7.7 7.68 5.8 5.8 7.65 7.64 s.d., 0.016 s.d., 0.091 Correlation, 0.76 Correlation, 0.90 5.75 7.6 7.6 5.75 7.6 7.64 7.68 7.72 7.76 7.8 0 200 400 600 800 1,000 5.75 5.8 5.85 5.9 5.95 0 200 400 600 800 1,000 State t Time, t State t Time, t Figure 1 | Some characteristic changes in non-equilibrium dynamics as a system approaches a catastrophic bifurcation (such as F 1 or F 2 , Box 1).
Box 2 j Critical slowing down: an example To see why the rate of recovery rate after a small perturbation will be reduced, and will approach zero when a system moves towards a � � catastrophic bifurcation point, consider the following simple dynamical � system, where c is a positive scaling factor and a and b are parameters: � d x which simplifies to ð 1 Þ d t ~ c ( x { a )( x { b ) � x 1 ) z d e x 1 ) z L f e [ d e It can easily be seen that this model has two equilibria, � x 1 5 a and x � f ( � d t ~ f ( � d t ~ l 1 e � L x x x 2 5 b , of which one is stable and the other is unstable. If the value of � � x 1 � parameter a equals that of b , the equilibria collide and exchange With eigenvalues l 1 and l 2 in this case, we have stability (in a transcritical bifurcation). Assuming that � x 1 is the stable x equilibrium, we can now study what happens if the state of the � l 1 ~ L f equilibrium is perturbed slightly ( x 5 � x 1 1 e ): x � ~{ c ( b { a ) � L x � d ( � x x 1 z e ) a ~ f ( � x x 1 z e ) and, for the other equilibrium d t Here f ( x ) is the right hand side of equation (1). Linearizing this equation � l 2 ~ L f using a first-order Taylor expansion yields � ~ c ( b { a ) � L x � � � b d ( � x x 1 z e ) x 1 ) z L f � ~ f ( � x x 1 z e ) < f ( � x e � � d t L x � � x 1 x � � If b . a then the first equilibrium has a negative eigenvalue, l 1 , and is thus stable (as the perturbation goes exponentially to zero; see equation (2)). It is easy to see from equations (3) and (4) that at the bifurcation ( b 5 a ) the recovery rates l 1 and l 2 are both zero and perturbations will not recover. Farther away from the bifurcation, the recovery rate in this model is linearly dependent on the size of the basin of attraction ( b 2 a ). For more realistic models, this is not necessarily true but the relation is still monotonic and is often nearly linear 16 .
Recommend
More recommend
Unleash a World of Digital Possibilities—Browse, Share, and Explore Content Without Boundaries