11/2/2011 Neuroscientists have been attracted to the puzzle of the - PDF document
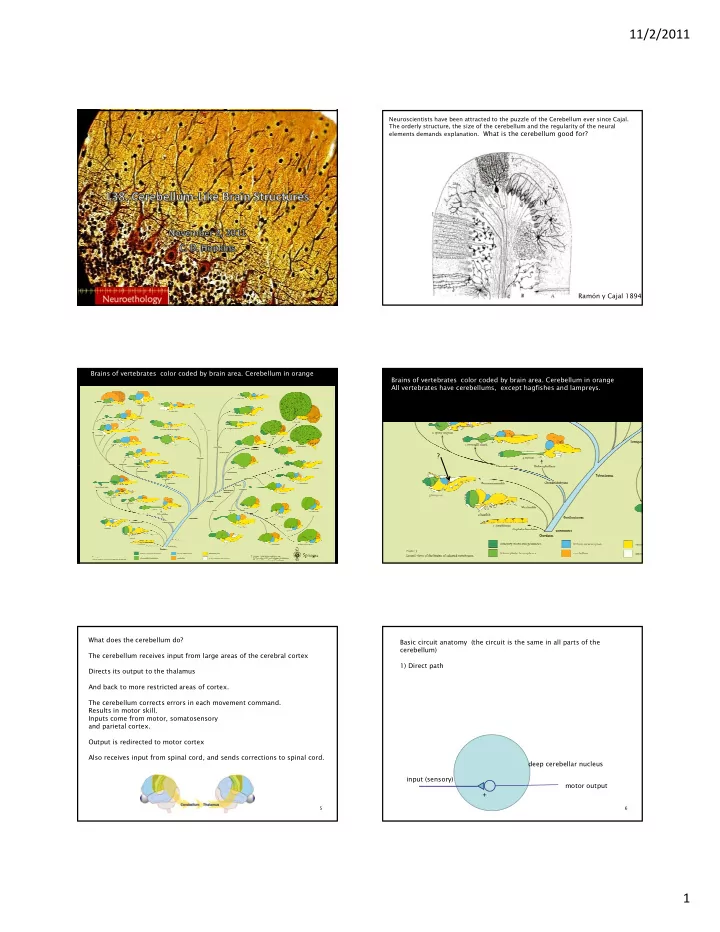
11/2/2011 Neuroscientists have been attracted to the puzzle of the Cerebellum ever since Cajal. The orderly structure, the size of the cerebellum and the regularity of the neural elements demands explanation. What is the cerebellum good for?
11/2/2011 Neuroscientists have been attracted to the puzzle of the Cerebellum ever since Cajal. The orderly structure, the size of the cerebellum and the regularity of the neural elements demands explanation. What is the cerebellum good for? Ramón y Cajal 1894 Brains of vertebrates color coded by brain area. Cerebellum in orange Brains of vertebrates color coded by brain area. Cerebellum in orange All vertebrates have cerebellums, except hagfishes and lampreys. ? What does the cerebellum do? Basic circuit anatomy (the circuit is the same in all parts of the cerebellum) The cerebellum receives input from large areas of the cerebral cortex 1) Direct path Directs its output to the thalamus And back to more restricted areas of cortex. The cerebellum corrects errors in each movement command. Results in motor skill. Inputs come from motor, somatosensory and parietal cortex. d l Output is redirected to motor cortex Also receives input from spinal cord, and sends corrections to spinal cord. deep cerebellar nucleus input (sensory) motor output + 5 6 1
11/2/2011 Basic circuit anatomy (the circuit is the same in all parts of the Basic circuit anatomy (the circuit is the same in all parts of the cerebellum) cerebellum) 1) Direct path 1) Direct path 2) Indirect path 2) Indirect path 3) Climbing fiber Purkinje Purkinje granule granule climbing fiber climbing fiber mossy mossy deep cerebellar nucleus deep cerebellar nucleus input (sensory) input (sensory) motor output motor output + + 7 8 When a nuclear stain (Nissl) is used, the density of granule cells is We recognize cerebellums from the cellular anatomy. They have: apparent. Purkinje cells are lightly stained, dendrites unstained. The Granule cells : receive input from mossy fibers , and send their outputs molecular layer looks mostly clear. to Purkinje cells via parallel fibers Purkinje cells : the “Principal Cell” of the Cerebellum Climbing fibers : bring “error signals” from the inferior olive to ONE Purkinje cell. 2
11/2/2011 Vestibular Ocular Reflex (VOR) + + Direct pathway from vestibular system to eye muscle Indirect pathway via cerebellum Signal from retinal “slip” inferior olive + + Marr, D. (1969). A theory of cerebellar cortex. J. Physiol. 202, 437–470. Albus, James A. (1971). A theory of cerebellar function. Math. Biosci. 10, 25– 61. “Influential theories of cerebellar function have posited that highly selective or sparse coding in Granule Cells allows Purkinje cells to acquire selective responses through associative synaptic plasticity” “Similarly for cerebellum-like circuits, such as the electrosensory lobe (ELL) of mormyrid fish, Granule Cells that selectively encode specific combinations of sensory and/or motor signals could allow Purkinje-like cells to generate more specific negative images.” “Nothing is yet known about how such signals are recoded in GCs or about the significance of GC input representations for the generation of negative images.” quotes from Sawtell (2010) Neuron. 3
11/2/2011 Learned changes in the VOR in monkeys outfitted Ito’s hypothesis with magnifying lenses. Visual input from superior colliculus to inferior olive: information about retinal ‘slip’ Climbing fiber input to Purkinje cell – error signal. Vestibular input from mossy fiber to granule cell to the parallel fiber in flocculo- nodular lobe of cerebellum. When co-active with error, synaptic weight of parallel fiber synapse is weakened by long term depression . Inhibition decreases and this strengthens the drive from the vestibular system increasing the gain. Cerebellum Function: DIRECT PATHWAY: the vestibular system to the eye muscles (VOR) The Basic Architecture INDIRECT PATHWAY: vestibular collateral (mossy fiber) � granule cell � parallel fiber � Purkinje cell (inhibits VN). ERROR SIGNAL: error in the VOR, information about image slip on the retina comes from the climbing fiber which originates in the inferior olive. (strong depolarization) Climbing fiber causes dendritic spike in the Purkinje cell � s causes synaptic weights from parallel fibers to depress, weakening the inhibitory input on the vestibular neurons, thereby adjusting the gain of the eye reflex. 1. Bell, C. C., Han, V. and Sawtell, N. B. (2008). Cerebellum-like structures and their implications for cerebellar function. Annu Rev Neurosci 31, 1-24. What is a Cerebellum-like structure? The cerebellar-like structure is defined by the molecular layer, the principal cell, and the numerous inputs from granule cells via parallel fibers. structure. Most vertebrates possess both a cerebellum and other brain structures with cerebellum-like architectures. 1) A MOLECULAR LAYER with many molec parallel fibers + dendrites of Purkinje-like cells. deep 2) A LARGE NUMBER OF GRANULE CELLS: which carry information from many central structures about the state of the central structures about the state of the animal: sensory information from a large number of senses, corollary discharges from motor centers (predictive of sensory input to the principle cells). 3) PRINCIPAL CELLS with SPINES. Parallel fibers terminate on spines of principal cells; or on the smooth dendrites of inhibitory stellate cells. Cerebellum-like structures sometimes serve as adaptive sensory filters that predict sensory inputs into the deep layers using the associated parallel fiber inputs in the molecular layer. 4
11/2/2011 Cunningham's Textbook of anatomy, by Daniel t b D i l John Cunningham, published in 1913 by William Wood Dendritic spines 25 26 The Basic Architecture of Mormyrid ELL 1. Bell, C. C., Han, V. and Sawtell, N. B. (2008). Cerebellum-like structures and their implications for cerebellar function. Annu Rev N i 31 1 24 How does the synaptic strength change with coincident inputs? 30 5
11/2/2011 Anti-hebbian. If the broad spike occurs after the EPSP, the synaptic weight is decreased. If the broad spike occurs before the EPSP, it strengthens. This sculpts a negative image of the expected input. broad spike before epsp broad spike after epsp acausal causal amplitude + change in epsp a - 500 ms + 500 ms - T (post-pre) delay time from epsp to broad spike Percentage change in excitatory postsynaptic potential (EPSP) amplitude plotted against the delay between EPSP onset and the broad spike peak during pairing. A negative delay with regard to the previous spike and a positive delay with regard to the following spike were present for each pairing. The shorter of these two delays is plotted. Filled circles, significant changes; open circles, non-significant changes (at P <0.01). 32 animation rule Gnathonemus petersii occurs when pre and post synaptic cells fire together 33 34 Natalia Caporale and Yang Dan (2008) Ann. Rev. Neurosci. Other cerebellar like structures show anti-hebbian plasticity according to slightly different learning rules. Tim Requarth and Nathaniel B Sawtell (2011) Curr. Opin. Neurobio. 35 36 6
11/2/2011 Formation of a negative image in the ELL is just one more mechanism for removing expected inputs from sensory event. • Adaptation of receptors or neurons to maintained stimuli removes responses to constant stimulus. • L t Lateral inhibition: remove expected mean levels l i hibiti t d l l over space.” Both of these methods use simple, non-plastic cellular processes (self inhibition, intra-cellular inhibition). 37 7
Recommend























More recommend
Explore More Topics
Stay informed with curated content and fresh updates.