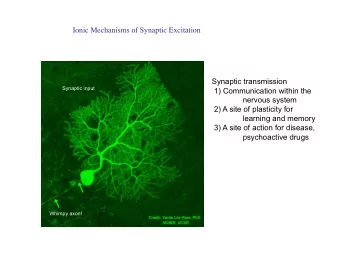
Foundations I Fall, 2016 Synaptic Transmission I
Neuromuscular Junction
Neuromuscular Junction junctional folds
Neuromuscular Junction
Transmitter Release Del Castillo and Katz (1954) m 0.9 mM Ca++ and 14 mM Mg++
Transmitter Release Fatt and Katz and Del Castillo and Katz (1951, 1952, 1954) Early experiments revealed that mEPPs could not result from the effects of a single molecule of ACh since iontophoresis or bath experiments with small concentrations (many molecules) did not give a discernible response. Thus, the mEPPs must result from the action of thousands of ACh molecules. So what controls the size of the mEPPs? How does the transmitter get released? Is it expelled in a continuous, graded fashion or is it constrained to some sorts of units (quanta), like energy? A statistical treatment was necessary since this is a phenomenon (mEPP size) that varied in amplitude and frequency. Del Castillo and Katz assumed that at the neuromuscular junction there was a large population of n units of undetermined nature that respond to a nerve impulse. To make the mEPPs as small as possible, [Ca2+] e was greatly lowered, Mg2+ was raised and a blocker of ACh receptors was added.
Transmitter Release Del Castillo and Katz (1954) If the probability of a single unit responding is p, and if each unit has an independent and equal p, then the mean number of units responding to each stimulus is given by: total n of available units mean quantal content m = np p of a unit responding Under these conditions, the relative occurrence of multiple events (i.e., mEPPs of sizes corresponding to integral multiples of the minimum mEPP size) is given by the Binomial distribution: n ! p x q ( n − x ) n x = N ( n − x )! x ! where N = the number of trials, nx is the number of events consisting of x units (quanta) and q = 1-p., i.e., the probability of the unit not responding.
Transmitter Release What is the probability of getting a response of size (number) x? x = n x P N substituting for n from last slide x n ! ( n − x ) x q P ( n − x )! x ! p x = But there is a problem calculating this...
Transmitter Release Del Castillo and Katz (1950s) ... the values of n and p are unknown! If one assumes that p is low (<0.1 or so), and equal for all n and that n is very large, the binomial distribution approaches the Poisson distribution described as: x = m x x ! e − m P m can be easily estimated in 2 ways: 1. m= mean amplitude of synaptic potential/mean amplitude of minimal synaptic potential 2. If one only counts the failures (x=0, mEPP amplitude = 0), the Poisson distribution degenerates to − m n 0 = Ne e − m = N m = ln N divide by N and take ln integrate n 0 n 0
mepp amplitude distribution poisson distribution The fit between the experimental data and the Poisson distribution was excellent. Thus, neurotransmitters are released in small packets called vesicles 1 vesicle= 1 quantum = Nobel Prize (Fatt, 1970)
But this is at the neuromuscular junction, not a synapse... ... and this was under stringent conditions that strongly interfered with transmitter release presynaptically... ... not to mention pharmacological blockade of post-junctional ACh receptors ... So how closely does this describe “normal” synaptic transmission? How many quanta are released at one time?
Mauthner interneuron AD spike and collateral IPSP Korn, Triller and Faber, 1982 stimulate intracellularly with HRP-filled microelectrode in presynaptic neuron record intracellularly from postsynaptic neuron label presynaptic neuron after experiment and count number of boutons
Mauthner Cell n is very small and p is huge! Korn et al., 1982
Korn et al., 1982
Korn et al., 1982 Transmitter release at this synapse is always monoquantal i.e., the presynaptic boutons operate in an all or none binary fashion!!!
Parabola: y=Ax-Bx 2 Y = PSC variance and x = PSC mean A and B are adjusted to fit parabola then Mean quantal size (Q) = A/(1+CV 2 ) Probability of release Pr= x(B/A)(1+CV 2 ) Number of release sites N= 1/B (curvature of parabola)
A2 A1 -47 mV -48 mV Traces 1-10 Traces 11-20 Traces 21-40 1.5 mV 0.2 mV 20 ms FS -->Spiny IPSP Spiny --> Spiny IPSP -18 pA -71 pA Postsynaptic Spiny neuron Postsynaptic Spiny neuron B1 B2 25 pA -136 pA 100 pA 100 ms 25 mV 25 ms Presynaptic FS interneuron Presynaptic Spiny neuron Tepper et al., 2004 Koós et al., 2004
cerebellar interneuronal IPSCs in vitro from verified single release site Auger et al., 1998
Auger et al., 1998 up to 30% of release events at this cerebellar synapse are multiquantal
Central Synaptic Transmission Sir Charles Sherrington (1857-1952) “In view therefore, of the probable importance physiologically of this mode of nexus between neurone and neurone it is convenient to have a term for it. The term introduced has been synapse.” - C.S. Sherrington, 1906
synaptic vesicle
Steps in Synaptic Transmission 1. A presynaptic impulse invades the terminal bouton and depolarizes it 2. The depolarization opens voltage sensitive Ca++ channels leading to a transient influx of Ca++ 3. [Ca]i triggers a sequence of biochemical events resulting in the fusion of a synaptic vesicle membrane with the terminal membrane.
Steps in Synaptic Transmission 4. The vesicle contents are extruded into the synaptic cleft in a process called exocytosis.
Exocytosis
Steps in Synaptic Transmission 4a. The vesicle membrane is incorporated into the presynaptic terminal membrane within ~ 50 µ sec of fusion. Excess membrane and vesicle components are recycled by endocytosis at sites outside the active zone into clatherin-coated vesicles that eventually lose the coating and are re-used 5. [Ca]i is immediately deactivated by uptake into mitochondria and presynaptic vesicles. 6. Released transmitter diffuses across synaptic cleft (very fast, no more than a few tens of microseconds). 7. Transmitter combines with postsynaptic receptors. 8. Receptor linked with ion channels and/or intracellular second messengers is activated and alters the permeability to certain ions and/or or cause an intracellular biochemical event
Synaptic Transmission II Voltage Clamp slope=I/V R=V/I R= 1/Slope Most synaptic potentials are due to an increase in conductance to an ion or ions
Synaptic Transmission II The increase in conductance causes a synaptic current to flow across the membrane I syn =n γ (E m -E syn ) where γ is the single channel conductance and n is the number of channels open. Recall that (Em-Esyn) is called the driving force The synaptic current is dependent on the driving force on the ions which depends on the potential difference between the membrane potential and the reversal potential. The point at which the driving force is zero is called the reversal potential. This is just another way of saying that the reversal potential is that membrane potential at which there is no net current flow.
Synaptic Transmission II a brief but necessary aside What is the difference between a reversal potential and an equilibrium potential? - Cl- free + HCO 3 Text - HCO 3 free - Cl- free + HCO 3 Kaila et al., 1989 Bicarbonate depolarizes the GABA equilibrium potential by up to A 10 mV depending on pH.
Synaptic Transmission II What does I syn =n γ (E m -E syn ) imply about the relation between membrane potential and synaptic potentials? Shifting the membrane potential (usually by injecting current through the recording electrode) must alter the driving force, thus altering the synaptic current thereby altering the amplitude of the synaptic response.
Synaptic Transmission II IPSP amplitude (mV) reversal potential mV mV E m (mV) mV mV mV stim
Synaptic Transmission II Voltage Clamp What is the reversal potential for the action of baclofen? I syn =n γ (E m -E syn ) reversal potential Because at that point there is no current flow due to the synapse, i.e., the control and the baclofen curves intersect. This can only happen when Em=Esyn Lacey et al., 1987
What does baclofen do to the membrane potential of the cell? What is this point? Em after baclofen What is this point? Em baclofen hyperpolarizes the cell by about 20 mV
Synaptic Transmission II A standard method to test for the synaptic nature of a membrane potential change (and to measure the reversal potential) is to inject current and alter the membrane potential and see the effect on the amplitude of the response.
Synaptic Transmission II Em Hyperpolarizing the membrane makes an EPSP get larger and and IPSP get smaller Depolarizing the membrane makes an EPSP get smaller and an IPSP get larger
Synaptic Transmission II Synaptic potentials that make the postsynaptic neuron more likely to fire an action potential are termed excitatory postsynaptic potentials (EPSP) and those that reduce the probability of the postsynaptic neuron firing are termed inhibitory postsynaptic potentials (IPSP). Note that this means that not all depolarizing synaptic potentials (DPSPs) are EPSPs.
Recommend
More recommend
Unleash a World of Digital Possibilities—Browse, Share, and Explore Content Without Boundaries